



海藻には紅藻、緑藻、褐藻などが存在するが、これら大型藻類は主に藻体の色の違いによって区別されることが多い。一方、海藻の「形」は多種多様で、種レベルでそれぞれ固有の「形」を持っている。したがって、「形」を参考にして種の見分けも大まかには可能である。例えば、私達が研究試料として扱ったことのある緑藻ミル属8種(図1)は外観の「形」を見れば種の同定が容易にできる。「形」から種名を言い当てられるのは他の海藻でも同様の場合が多い。では、この海藻の「形」はどのように作られ、決定づけられているのか?日頃から知りたいと思っていた。陸上植物にも様々な「形」が存在するが、その「形」は内在する形態形成関連遺伝子を基礎として、植物ホルモン等による種に固有の遺伝子発現制御の結果として作られるものと考えられる。このエッセイ執筆を担当することになったことを良き機会として、海藻の形態形成はどうなっているのか、関連研究の進捗状況はどうなのか、文献を中心に調べてみることにした。

海藻では、陸上植物に見られる根、茎、葉の器官の分化は見られず、体全体が葉状であり、この葉状体(広義の)が外観となる。この葉状体には糸状(ジュズモ、シオグサ、モズクなど)、膜状(狭義の葉状体)(マキヒトエ、アオサなど)、管状(ミル、ハネモなど)などの形態が存在する。一方、異形世代交代をもつアマノリ類のように、同一種にもかかわらず糸状(胞子体、2n)と膜状(配偶体、n)の2種類の明らかに異なる形態をもつものも存在する。アマノリ類の場合は便宜的に前者を糸状体、後者を葉状体(狭義の)と呼んで区別している。上述の各葉状にはさらに種に依存した多様な「形」が存在する。
これら広義の葉状体が海藻の「形」に相当するわけであるが、それはどのように形成されるのであろうか。この海藻の形態形成に関して、とても興味深い研究例がある。主に20世紀後半から今世紀初頭にかけて、複数の藻類研究グループによって、海藻を無菌で培養すると正常な形にならず不定形の細胞塊(カルス)状になるが、海藻表面から分離した細菌や細菌培養液の存在下では正常な葉状体が誘導されることが見出された。研究に用いられた海藻は主としてマキヒトエ(Gayralia oxysperma)やアオノリ類が属するマキヒトエ属・アオサ属緑藻であるが、紅藻、褐藻でも同様の無菌培養によるカルス状の形態形成が確認されている(表1)。これらの研究の多くは日本人研究者によってなされているが、さらに2003年に松尾らは海藻や海綿から単離した1000株以上の付着細菌について緑藻マキヒトエに対する形態形成誘導活性を検索し、正常葉状体

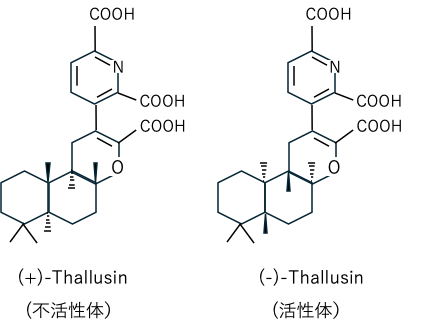
への形態誘導活性を示す50種類の細菌株を見出した。その多くはCytophaga-Flavobacterium-Bacteroides (CFB) グループ内のグラム陰性細菌Zobellia uliginosaのクレード(分岐群)に分類されることもつきとめた。また、これら細菌は他のアオサ属緑藻の無菌胞子に対しても正常形態形成を誘導すること、マキヒトエ属緑藻の若い葉状配偶体からの胞子放出活性をもつことも明らかにした。極めつきは、マキヒトエグサ属緑藻から単離した表面細菌株 YM2-23の培養液140 L(1L容フラスコ x 175個)から極微量(140 µg)のサルーシン (thallusin; thallus inducing factor)と名付けた形態形成物質を単離し、その構造を明らかにしたことである(図2)(Matsuo et al. 2005)。サルーシンのマキヒトエに対する最小有効濃度は 1 fg/mL~1 ag/mL(10-15~10-18g/mL)で極めて微量で活性を示すこと、この物質は供試した他のアオサ属緑藻にも有効であることが示された。一方、サルーシンの添加を止めると葉状体の分解が起こることから、自然環境下ではサルーシン含有細菌が常時マキヒトエ表面に付着していることが正常な形態形成の維持に必要であることが示された。本研究成果は藻類学における快挙であり、海藻付着細菌と海藻との相互作用(共生)関係の大切さを示唆している。現在、本研究成果の応用として、産官学の協働下で海藻付着細菌やサルーシンを用いたアオノリ類の陸上養殖も試みられている(Wichard 2023、木下 2024)。応用に際しては、サルーシンの細菌からの供給に量的制約があることから、その化学合成研究も行われ、合成された計28種類のサルーシン類似体の構造活性相関を調べるとともに、最終的に不斉全合成にも成功して図2に示す絶対立体配置 ((-)-Thallusin)が決定された。しかし、サルーシンが形態形成を誘導するメカニズムや細菌内でのサルーシンの生合成系は未知で今後の興味ある重要課題となっている。
付着細菌の形態形成能については、紅藻スサビノリ(Neopyropia yezoensis)においても調べられている。嵯峨らのグループはスサビノリを無菌下で培養すると、配偶体世代において葉状の生長や正常な形態形成が欠損し、カルス状の形態になることを認めるとともに、スサビノリ表面から分離した共生細菌2株(BPY-W6とBPY-W8)の存在下で培養すると正常の葉状体(配偶体)に生長することを明らかにした。また、スサビノリ由来プロトプラストも無菌培養下ではカルス状藻体となるが、グラム陰性細菌に属するヒフォモナス属細菌(α-proteobacteria)を添加すると正常な葉状体に生長することも報告されている。このヒフォモナス属細菌株はスサビノリ仮根に特異的に結合することにより形態形成を誘導するようである。しかし、スサビノリに関しては、生長・形態を誘導する共生細菌の存在は明らかにされたが、それらが作り出す形態形成物質の同定には至っていない。今後、紅藻や褐藻の形態形成に機能する共生細菌由来の形態形成物質の発見が待たれる。他方、植物ホルモンのオーキシン(インドール3-酢酸)がナラワスサビノリ(Neopyropia yezoensis f. narawaensis)において頂茎細胞の再生や生長を促進することが明らかにされている。これは従来から知られる植物ホルモンが紅藻の生長、分化にも関与することが明らかにされた最初の例と思われる。しかし、陸上植物に存在するオーキシン受容体遺伝子はナラワスサビノリには存在しないことから、海藻(ナラワスサビノリ)においては陸上植物とは異なる植物ホルモンの作用機構が存在すると推定されている。
これまで述べたように、ある種の海藻においては正常な葉状体への形態形成が藻体表面の着生細菌によって制御されていることは理解できた。しかし、海藻における種に固有の「形」作りの詳細は依然として謎である。