



その頃,筆者はこの課題を解決するためどのような研究をすれば良いのか,すなわち形態や生活史型以外の特徴で褐藻の系統関係を明らかにする方法がないか,いろいろ試行錯誤していた。その候補の一つとして考えたのが,当時,教科書などでも紹介されはじめていた分子進化学的な手法であった。化石がほとんど産出しない褐藻の研究で,遺伝子やタンパク質の情報に基づき系統上の関係を推定し,また分子時計とよばれる手法でその進化年代を推定できる研究はきわめて魅力的であった。しかし,筆者が研究を始めた頃は,海藻の系統関係を明らかにすることを目的とした分子生物学的な解析は,塩基配列のみならず酵素やアミノ酸配列を用いたものを含めて全く行われていなかった。このため,1980年代半ばには5SリボソームRNA配列による分子系統の研究を行っておられた堀寛氏に共同研究をお願いして,コンブモドキを含むいくつかの褐藻と不等毛植物の解析を行い,その結果が生活史の論文と同じ1986年に出版された。しかし,あいにくわずか約120塩基対のこの分子種では「褐藻類の誕生がおおよそ2億年前で,10億年を超える紅藻よりはるかに新しい」という推定はできたが,コンブモドキと他の褐藻の関係という細かい議論は全くできなかった。
![図6 コンブモドキを含む分子系統樹。葉緑体ルビスコ遺伝子系統樹 (ML) [Sasaki et al. (2001) を改変].](img/photo_039_07.jpg)
[Sasaki et al. (2001) を改変].
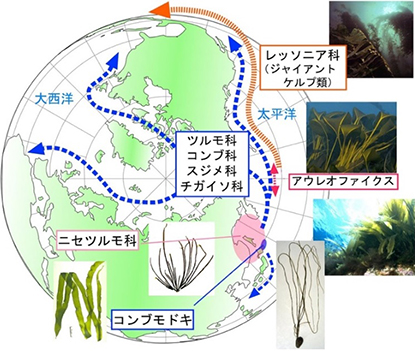
その後,サンガー法によるDNA塩基配列解析法やPCR法の普及によって,自らの研究室でも分子系統学的研究が行えるようになった。そこで,筆者は加藤敦之氏の協力を得て,韓国の研究グループと共同で核の18SリボソームDNA配列を用いたコンブ類の系統解析を行い,ニセツルモとツルモが近縁であり,また両者が他のコンブ類と近縁であることを1999年に報告した。そして,2001年には核リボソームDNAの介在領域や葉緑体ルビスコ遺伝子の塩基配列による解析で,コンブモドキがニセツルモと近縁で,また両者がツルモを含むコンブ目と最も近縁であることを示し,コンブモドキを独立した科,コンブモドキ科としてコンブ目に含めることを提案した(図6)。また,これらの系統関係をふまえて,いわゆるコンブ類が太平洋から大西洋,また北半球から南半球に至る広い海域に分布するのに対して,コンブモドキやニセツルモは太平洋北西岸の北東アジアだけにしか分布しないことから,コンブの祖先はこのあたりで誕生し,ベーリング海を経て広く世界に広がったのでは,という分布拡大のストーリーも描くことができた(図7)。
さて,この段階で「コンブモドキ」は「コンブ」なのか,との長年にわたる疑問にようやく答えをだすことができた(やはり「コンブモドキ」は祖先的な「コンブ」だった・・・)と思ったのだが,そうは問屋が卸さなかった。その当時の分子系統解析は,ある特定の遺伝子の塩基配列情報に基づき分子系統樹を構築し,それぞれの分岐の系統関係の確かさをブートストラップ法などで統計学的に検証するのが一般的な手法だった。しかしながら,どの遺伝子による解析でも大形で複雑な形態をしたコンブ類のまとまりは統計的に強く支持されるのに対して,ツルモ,コンブモドキ,ニセツルモと他のコンブ類の関係は,この3者がひとかたまりのグループになることは間違いない(すなわち,別の褐藻のグループが3者の間に入ることはあまりない)のだが,その支持はあまり高くならない。そこで,2013年には葉緑体やミトコンドリアの8つの遺伝子の連結配列による分子系統解析を行ったが,状況はあまり改善しなかった(図8)。さらにその後,カナダのグループに協力して行ったいわゆる次世代シークエンスによる数十を超える遺伝子に基づく最新の多遺伝子系統解析でも,ツルモ,コンブモドキ,ニセツルモの系統上の近さは確認されたが,「コンブ」とはやはり近いものの,一つのまとまったグループというほどの根拠は示せなかった。そして,2019年に出版された論文では,これらの結果と共同研究者の意向もふまえて,ツルモとコンブモドキ,ニセツルモはコンブ目から外して,ツルモ目という新しいグループとして分類することを提案することになった(図9)。
![図8 コンブモドキを含む分子系統樹。葉緑体・ミトコンドリア8遺伝子系統樹 (MP)[Kawai et al. (2013) を改変].](img/photo_039_09.jpg)
[Kawai et al. (2013) を改変].
![図9 コンブモドキを含む分子系統樹。葉緑体・ミトコンドリア・核多遺伝子系統樹 (ML) [Starko et al. (2019)を改変].](img/photo_039_10.jpg)
[Starko et al. (2019)を改変].
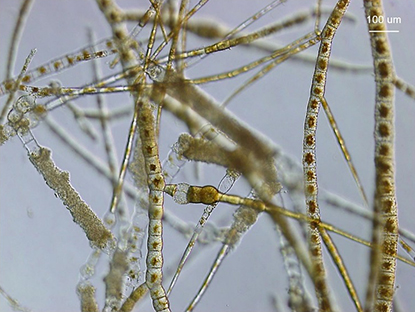
一方,2014年にシルバーフェルト (T. Silberfeld) らは,1988年にフーパー (R.G. Hooper)らによってカナダ大西洋沿岸からチロプテリス目の種として記載されたフェオシフォニエラ (Phaeosiphoniella cryophila) が,葉緑体とミトコンドリアの7遺伝子に基づく分子系統解析ではコンブモドキ,ツルモを含むコンブ目と近縁だが,独立した目として分類するとの論文を発表した。しかしこの海藻の藻体は分枝した糸状で,形態上はコンブ類のみならずコンブモドキとも大きく異なる(図10)。あいにく,前述の最新の多遺伝子解析ではフェオシフォニエラは加えられなかったため,改めてこの種を加えた多遺伝子解析を進めているが,これもやや予想外の結果になりそうである。
この様に,筆者の35年におよぶ「コンブモドキ」は「コンブ」なのか,という問いはいまだに決着せず,むしろ混迷を極めている。このため,一研究者としては新たな展開にわくわくする一方で,定年を過ぎた身としては,さていつになったら片付くのか,ついつい憂鬱になってしまうのである。
執筆者
川井浩史(かわい・ひろし)
神戸大学内海域環境教育研究センター特命教授(同元センター長)・名誉教授、理学博士、日本藻類学会元会長、アジア太平洋藻類学会 (APPA) 元会長、日本藻類学会学術賞 (山田賞) 受賞(2019)
「「コンブモドキ」は「コンブ」か —終わりの見えない問い— 2/2/川井浩史 | 海苔百景 リレーエッセイ」ページのトップに戻る